
From the Department of Physical Medicine and Rehabilitation, University of Texas Health Science Center at Houston McGovern Medical School and TIRR Memorial Hermann, Houston, Texas, USA
Objective: Botulinum neurotoxin is commonly utilized in neurorehabilitation as a treatment for focal spasticity. Clinical experience has yielded observations of late motor recovery after intramuscular injection of botulinum neurotoxin, that are not readily explained by the classical mechanism of action of the neurotoxin in controlling spasticity. These findings have triggered speculation regarding a botulinum neurotoxin mediated effect at the central level after peripheral intervention.
Methods: A review of current literature reveals evidence of distant action after peripheral botulinum neurotoxin injection in affected muscles, be it in other muscles, nerves, spinal cord or the cortex.
Results: Plausible explanations for a centrally mediated late motor recovery after botulinum neurotoxin injection include: (i) direct action of botulinum neurotoxin at distant sites in the central nervous system, mediated by retrograde transport of the neurotoxin into the spinal cord, and (ii) cortical reorganization due to botulinum neurotoxin-induced decrease in peripheral sensory input at the local injection site.
Conclusion: Additional research is required to further elucidate these hypotheses, as well as providing specific dosing specifications, patient selection criteria and the interplay with other therapeutic modalities necessary to promote late motor recovery.
Key words: botulinum toxin; plasticity; motor recovery.
Accepted May 23, 2017; Epub ahead of print Aug 9, 2017
J Rehabil Med 2017; 49: 00–00
Correspondence address: Manuel F. Mas, 1333 Moursund Street, Houston, Texas, USA, 77030. E-mail: manuel.mas@uth.tmc.edu
Botulinum neurotoxins (BoNTs) are the most poisonous biological substances known, and are produced by anaerobic bacteria of the genus Clostridium (1, 2). Despite this, BoNTs are used as treatment options for numerous ailments in the medical field. Chemodenervation with BoNT is a commonly used intervention for the treatment of spasticity and other components of upper motor neurone syndrome (3). Treatment goals include reducing tone and pain, increasing range of motion, and improving ambulation and hygiene. It is widely accepted that the mechanism of action of BoNT occurs in the peripheral nervous system through the blockade of acetylcholine release at the neuromuscular junction. Recently, central effects of BoNT are suspected due to electrophysiological changes in both distant muscles and nerves. Evidence of local and systemic analgesic effects of BoNT/A has been postulated in rat models for polyneuropathy, decreasing pain in the side contralateral to the injection (4, 5). Distant and contralateral muscles have been shown to be inhibited after BoNT/A injection of a different muscle (6–10). More so, reversible changes in cortical organization and activation have been noted in advanced imaging after injecting BoNT/A peripherally (11, 12). These clinical and experimental findings raise the question as to whether BoNT is able to produce effects at the spinal and supraspinal levels and, if so, through what mechanism?
Our clinical practice has provided clinical examples of improvements in areas other than spasticity after targeted BoNT injection in overactive muscles. These favourable outcomes are not readily explained by the peripheral mechanism of action of the neurotoxin, but by a potential effect at the central level. This review article uses these cases as a platform for discussion of the current evidence regarding a possible central effect of BoNT, and to hypothesize potential mechanisms of action for unanticipated robust functional outcomes post-BoNT treatment.
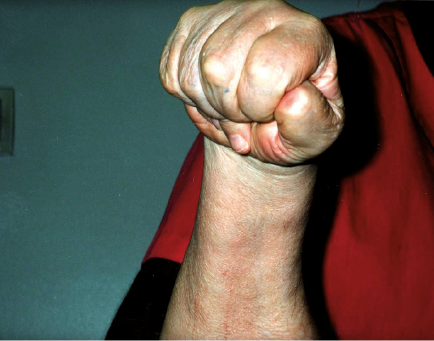
A 67-year-old man requested BoNT injection to recover the use of his right hand approximately 4 years after having a traumatic and hypoxic brain injury and brainstem stroke following a motor vehicle crash. Brain imaging data was not available. He presented with right hemiparesis and a clenched fist (Fig. 1). Modified Ashworth Scale (MAS) score was graded as 4 in the right flexor digitorium superficialis (FDP), flexor digitorium profundus (FDP), flexor pollicis longus (FPL), flexor pollicis brevis (FPB) and lumbricals. MAS was one in the right forearm pronators and elbow flexors. There was no clinically detectable active finger movement on evaluation. The clinician and the patient agreed that the goal of treatment was to increase finger range of motion to facilitate hygiene and prevent further contractures. A total of 200 units of onabotulinumtoxinA was administered using ultrasound guidance (100 units each to the right FDS and FDP, 2 injections sites per muscle). Four weeks post-injection, the patient’s previously clenched fist opened due to relaxation of the finger flexors (Fig. 2). Moreover, the patient demonstrated active, albeit limited, finger flexion and extension. Metacarpophalangeal flexion persisted. Approximately 3 months post-injection, the patient had another onabotulinumtoxinA injection (250 U total) distributed to the right FDS (100 units in 2 sites) and FDP (100 units in 2 sites), FPL (10 units) and lumbricals (10 units to each, 40 units total) with ultrasound guidance. Approximately one month later, the patient recovered more active finger flexion and extension (Fig. 3) and entered a 5-day-a-week occupational therapy programme focusing on “forced-use” of the right upper limb. Approximately another month later, the patient recovered further active movement of the right fingers that allowed him to button and unbutton his shirt, wipe his mouth with a tissue, and use a hairbrush without adaptation.
Fig. 1. Case 1: initial presentation.
Fig. 2. Case 1: 4 weeks after first injection displaying relaxation of finger flexors.
Fig. 3. Case 1: 1 month after second intervention. Increased recovery of active finger flexion and extension.
A 74-year-old woman presented to the spasticity management clinic for BoNT injection. She visited our clinic 38 years after an ischaemic stroke with residual left hemiparesis and a clenched fist due to chronic spasticity (Fig. 4). There was no clinically detectable active finger movement on evaluation. MAS score was 4 at FDS, FDP, FPL and FPB. The treatment goal was to facilitate cleaning of the palm of the hand, which had become difficult over the years. A total of 50 units of onabotulinumtoxinA were injected using ultrasound guidance (25 units to the left FDS in 2 sites, 15 units to the left FPL and 10 units to the left first lumbrical). Four weeks after injection, she reported no clear benefit from the intervention. She had a second course of onabotulinumtoxinA injections (100 units total) to the left FDS (50 units in 2 sites), left FPB (20 units), and left first and second lumbricals (15 units each) with ultrasound guidance. Three months later, the patient reported greater relaxation yet still having difficulty with releasing grip. She agreed to another course of onabotulinumtoxinA injections with ultrasound guidance, 200 units total, distributed to the left FDS (100 units in 2 sites), left FDP (45 units in 2 sites), FPB (25 units) and left first and second lumbricals (15 units each). At follow-up evaluation 3 months later she was pleased to report greater relaxation of the fist and, more surprisingly, active limited finger flexion and extension for the first time since the onset of stroke 38 years earlier. Of note, she did not perform any exercise in the interim.
Fig. 4. Case 2: initial presentation.
Classically, BoNT has been utilized in rehabilitative treatment as a tool to reduce spasticity. This has been well documented in noteworthy randomized controlled trials, proving its efficacy for both upper and lower limb spasticity (13, 14). This has been attributed to the action of the toxin in the periphery, at the neuromuscular junction.
BoNT targets the neuromuscular junction, where it blocks acetylcholine release, presynaptically producing partial paralysis. It enters the cell where it cleaves the SNARE protein complex by way of its metalloproteolytic activity, preventing exocytosis and neurotransmitter release (15, 16). Its effects are dose-dependent and reversible (17). This enzymatic action is directed at the distinct neuronal components of the neuromuscular junction and reflex arc. Skeletal muscle contains extrafusal and intrafusal muscle fibres, which are responsible for muscle contraction, and sensory and proprioceptive feedback, respectively. Extrafusal fibres are innervated by alpha motor neurones, comprising a motor unit. The connection between the extrafusal fibre and a single alpha motor neurone nerve ending is known as a neuromuscular junction. On the other hand, gamma motor neurones innervate intrafusal fibres, forming the muscle spindle serving as mechanoreceptors for the muscle and providing feedback to the central nervous system. The decrease in excessive muscle tone caused by the targeted effect of BoNT in alpha motor neurones at the neuromuscular junction is the primary endpoint for clinicians treating spasticity. BoNT is also reported to cause reflex inhibition by blocking intrafusal muscle fibres with consecutive reduction of Ia/II afferent signals and muscle tone without affecting muscle strength (18).
BoNT has been postulated to possess analgesic effects (19). It has been studied and, at times, used for the treatment of migraine (20–22), arthritic joint pain (23, 24), myofascial pain syndrome (25) and trigeminal neuralgia (26, 27). Recent literature also points to pain reduction after local BoNT injection for spasticity secondary to numerous upper motor neurone syndromes, including stroke (28, 29). Local analgesia can be the result of local neurotransmitter inhibition from sensory nerve endings by cleavage of peripheral synaptosome associated protein 25 (SNAP-25) (30). BoNT can also directly prevent release of glutamate, calcitonin-gene related peptide, substance P, peripheral sensitization, formalin-induced pain and expression of transient receptor potential vanilloid, all mechanisms of peripheral pain generation (19). Yet, analgesic effects via BoNT remain a matter of debate, requiring more research and scrutiny of the available data.
Adverse clinical effects following intramuscular injection of BoNT are well established, with some patients reporting muscle weakness, oropharyngeal/respiratory alterations, bowel and bladder disturbances and infections, some of which require higher levels of care (31). Instrumented guidance of BoNT injection, such as electrical stimulation or ultrasonography, is strongly recommended (32, 33). Guided injections may improve outcomes (34) and decrease adverse events due to increased accuracy.
Recently, BoNT-mediated effects on other components of the neuromuscular junction, such as gamma motor neurones and the muscle spindle/intrafusal fibres, have been studied further, as noted in its potential central modulation of pain (19) and distant effects on non-targeted muscles (9). BoNT has been postulated to act at the level of gamma motor neurones, decreasing Ia-afferent input to the central nervous system. This local effect of decreased sensory input induced BoNT has been utilized as a central theory behind much of the research, pointing at distant effects on components of the central and peripheral nervous system as well as on other muscles.
There is a considerable amount of information suggesting that BoNT has effects distant from the local injection site (5, 8). These include the alteration of muscle and nerve physiological properties distant from the intervention area. Unilateral injection of BoNT in the orbicularis oculi muscle caused contralateral alteration of electrophysiological parameters, permitting the possibility of toxin spreading outside the neuromuscular junction (6). Similar findings were echoed in a study evaluating sternocleidomastoid muscle parameters after unilateral BoNT injection, evidencing contralateral weakness after long-term treatment (7). Abnormal jitter has been documented in distant muscles after local BoNT injection, again alluding to a possible distant effect of the neurotoxin (8, 10).
In 2013, Marchand-Pauvert et al. (9) and others reported a reduction in posterior tibial nerve inhibition of vastus lateralis H-reflex after BoNT/A injection in the triceps surae of stroke patients. It was postulated that this reduction in spinal recurrent inhibition was induced by BoNT/A injected peripherally. It was hypothesized that distant muscle effects were caused by a modification of the reciprocal inhibitory pathway. As described by Renshaw in 1941 (35), alpha motor neurones that innervate the neuromuscular junction interact with Renshaw cells in the spinal cord. In turn, Renshaw cells mediate recurrent inhibition to other spinal motoneurons (35, 36) and are a potential target for retrogradely transported BoNT into the spinal cord. Marchand-Pauvert and colleagues (9) demonstrated that injection of BoNT/A into the posterior tibial nerve innervated triceps surae muscle “disinhibited” the vastus lateralis muscle in stroke patients, a muscle innervated by the femoral nerve. The group theorized that these findings were a result of BoNT/A affecting spinal synaptic transmission and reciprocal inhibition by way of retrograde transport of the neurotoxin and modulation of the activity of Renshaw cells. Aymard and colleagues (37) studied the effects of BoNT in reciprocal inhibition in patients with post-stroke spasticity. They proved that reciprocal inhibition of the tibialis anterior muscle mediated by the posterior tibialis nerve was altered after peripheral BoNT injection in the ankle plantarflexors. The group postulated that BoNT produced central action, modulating spinal plasticity by way of retrograde transport along motor axons to Renshaw cells. Others have also observed alteration in H-reflex parameters of antagonist muscles in different conditions after BoNT injection. Patients with focal upper limb dystonia displayed increased reciprocal inhibition of forearm flexor and extensor muscles after local BoNT/A injection (38). Similar results were obtained in a separate study in patients with essential hand tremor, pointing at restoration of presynaptic inhibition in forearm antagonist muscles after BoNT/A injection due to probable action of the toxin at both extrafusal and intrafusal muscle fibres, the latter resulting in decreased muscle spindle afferent input to the spinal cord (39). Wohlfarth and others (40) reported prolongation of F-wave latencies and reduction in F-wave persistence of the untreated abductor digiti minimi and tibialis anterior muscle after BoNT/A injection in patients with spasmodic torticollis and writer’s cramp.
Recent animal studies have also demonstrated analgesic effects in locations far from the initial injection site. Bach-Rojecky et al. (4) utilized a rat model for diabetic neuropathy to evaluate the effect of subcutaneous and intrathecal administration of BoNT/A. The results revealed bilateral pain reduction after unilateral toxin application, with faster onset from a lower intrathecal application compared with a higher subcutaneous dose. This was theorized to be a result of retrograde transport of BoNT into the central nervous system, specifically at the spinal cord level. This permits indirect action on different targets, a theory also proposed by others (21, 41). This body of evidence opens the door to the possibility of effects of BoNT outside of the neuromuscular junction and in the spinal cord.
The distant effects of BoNT have been observed even farther from the local injection site, at the supraspinal level. For instance, there is research that presents the possibility of BoNT-mediated induction of cortical reorganization. Evaluation of long-latency reflexes of hand muscles in BoNT/A-treated idiopathic focal dystonia has shown evidence of reduction in amplitude of cortically mediated long-latency reflexes after peripheral toxin injection (42). Somatosensory evoked potentials (SEPs) have also been used to evaluate cortical effects of peripheral BoNT application. Several studies have shown that cortical SEPs are altered after BoNT injection of affected muscles in different populations, perhaps demonstrating central reorganization after intervention secondary to modulation of spindle afferent inputs (43–45). However, the sensitivity of cortical SEPs to assess cortical sensory effects of peripheral BoNT interventions remains debatable (46).
Advances in imaging techniques have expanded the study of the cortical effects of BoNT. In 1998, Byrnes and others (47) used transcranial magnetic stimulation (TMS) to observe how patients diagnosed with writer’s cramp had corticomotor representation that differed from that of healthy subjects, with shape distortion and overextension of lateral borders. These abnormalities in the primary motor cortex were normalized after BoNT/A injection, resembling that of healthy individuals. These changes were reversed to baseline after the clinical effect of BoNT/A dissipated. Similarly, primary motor cortex excitability was analysed using TMS in BoNT-treated patients with cervical dystonia. After peripheral toxin injection, there was evidence suggestive of changes in plasticity in the primary motor cortex hand area potentially mediated by BoNT modulation of afferent inputs (48). TMS has also been used to analyse intracortical inhibition after BoNT injection. Several studies have reported how intracortical inhibition was normalized after peripheral injection of BoNT/A. Huynh et al. (49) observed how patients with post-stroke spasticity had an increase in short-interval intracortical inhibition in the contralesional hemisphere after peripheral injection of BoNT, normalizing to control values. This effect peaked at 7 weeks, correlating with clinical measures of spasticity and returned to pre-injection parameters at 11 weeks. The changes in clinical spasticity preceded the changes in intracortical inhibition. The group concluded that pre-existing maladaptive plastic responses could contribute to post-stroke spasticity and that BoNT can normalize these by several methods including: haematogenous dissemination, retrograde transport of the neurotoxin into motor cortices through motor fibres and propriospinal pathways, and/or action at the intrafusal neuromuscular junction by way of deafferentation. These findings were echoed by Gilio et al. (50) in patients with upper limb dystonia after local BoNT injection, again suggesting sensory afferent modulation of the central post-stroke physiology.
One of the most recently developed neuroimaging techniques, functional magnetic resonance imaging (fMRI), has also been utilized to study the central effects of peripheral BoNT. Cortical reorganization and deactivation of additional cortical areas recruited during impaired spastic hand movement has been evidenced using fMRI. In 2013, Tomasova et al. (11) utilized fMRI to evaluate cortical activity after BoNT/A injection of the spastic arm muscles of hemiplegic patients after stroke. Imaging was obtained before treatment, at weeks 4 and 11 after injection. Group imaging before injection revealed extensive participation of contralateral primary motor cortex, supplementary motor area, bilateral premotor cortices, superior parietal lobe and basal ganglia. This activation reduced significantly 4 weeks after injection, only to reappear grossly 3 months after intervention coinciding with the reappearance of spasticity. This BoNT-induced reduction in extent and lateralization of aberrant cortical activation closely resembled findings of motor recovery after stroke in past fMRI studies. It was proposed that cortical overactivation was a compensatory mechanism for motor output after stroke and could be modulated after BoNT injection. The group hypothesized that BoNT produced this cortical effect by blockage of the neuromuscular junction of gamma-motor neurones, reducing Ia afferent signals to the central nervous system, and echoing theories postulated by other studies mentioned previously. These findings coincide with the ones reported by Veverka et al. (12). However, the latter also found volume reduction in the frontoparietal sensorimotor network during the eleventh week after peripheral BoNT injection, well beyond the expected time of medication effect. This group concluded that BoNT could modify cerebral plasticity despite its temporary effect in the injected muscle. Recently, fMRI served to study cortical activation in both cerebral hemispheres after BoNT/A injection in stroke patients with right hand spasticity. Patients who had experienced stroke had significantly greater activation in motor and pre-motor cortex compared with healthy subjects, especially on the non-injured hemisphere. After injection, there was a normalization of cortical activation between both hemispheres with greater decrease in ipsilateral (non-injured) motor and pre-motor cortex (51), similar to the findings reported by others.
As discussed, there is considerable literature documenting the effects of BoNT far from its local target at the level of the muscle. These range from contralateral muscles, spinal cord and cortical regions. Yet, how do these findings translate to the clinical cases presented previously? Both patients actively flexed and extended their fingers after several peripheral BoNT/A injections, one of them without participating in any form of therapy. What are the mechanisms that allowed for BoNT to yield late motor recovery in both patients? There are several mechanisms that could allow BoNT to produce these favourable results. Some of these have been already discussed by some of the literature reviewed previously. We will examine these potential mechanisms of action and evaluate how these might explain our clinical findings.
Before acting at distant sites, BoNT requires a transport route. The circulatory system has been proposed as a possible carrier for BoNT to reach the central nervous system, by haematogenous dissemination through the blood-brain barrier (49). However, there is currently no evidence that peripheral therapeutic doses of BoNT can cross the blood-brain barrier (46). Retrograde axonal transport is another dissemination method that could potentially carry the toxin to the spinal cord sensory and motor neurones (52). Under resting conditions, BoNT/A has been postulated to bind to the fibroblast growth factor receptor 3 (FGFR3), which is expressed at motor nerve terminals and undergoes endocytosis (53). This pathway may direct the neurotoxin to an endosomal compartment from which BoNT/A enters axonal carriers targeted for retrograde transport (54). BoNT can also produce central effects without distant spreading, instead by modulating circuitry that reaches the central nervous system. In fact, this model has been proposed in the literature and mentioned earlier, constituting presynaptic blockade of the connection between gamma-motoneuronal endings and intrafusal muscle fibres (52). This can reduce input from group Ia afferent fibres and could alter spinal pathways excitability as well as motor maps at the cortical level, a phenomenon discussed previously (54).
Both clinical cases exhibited late spastic hemiplegia after their original central insults. BoNT has been well established as a therapeutic agent to decrease spasticity. Yet, it is the apparent modulation on weakness that raises the question of central action of the neurotoxin. Late motor recovery after peripheral BoNT/A treatment for spastic-plegic hand can be a result of modulation of dormant central circuitry controlling the immobile muscles, as well as their antagonists. It can also result from deactivation of central nervous system structures inhibiting motor output of the observed muscle groups; that is, by unmasking residual function. Modification of cortical activity or topography could potentially explain agonist and/or antagonist activation. A detailed study of the circuitry that generates motor activity in an extremity (e.g. the hand muscles) is beyond the scope of this work. Nevertheless, a general map is crucial to hypothesize potential locations of BoNT action to produce movement in the spastic-plegic muscles. Overall, movement originates in the cortex, where the primary motor cortex, supplementary motor area and premotor cortex project to the corticospinal tracts. Other areas that project to this pathway are the somatosensory cortex, parietal lobe and the cingulate gyrus. Axons project through the posterior limb of the internal capsule and into the brainstem and anterior medulla. Fibres later separate into the anterior and lateral corticospinal tracts in the spinal cord. Upper motor neurones then synapse with an interneurone and later with the lower motor neurone in the anterior horn of the spinal cord. Lower motor neurones, specifically alpha motor neurones, then innervate extrafusal muscle fibres and commence skeletal muscle contraction.
Using this framework, peripheral BoNT can modify one or several parts of this pathway to produce movement in a spastic-plegic hand. Previously cited studies have pointed at cortical reorganization after peripheral BoNT injection. Yet, these have not noted activation of previously silent points in cortical motor generation, but rather a decrease in aberrant over-stimulation of cortical topographical areas recruited in patients with spastic limbs. As discussed previously, Tomasova et al. (11) demonstrated how patients presenting with spastic hemiplegia who were treated with BoNT had fMRI changes in cortical activation with restriction to midline and contralateral sensory and motor cortices, following a reduction in extent of cortical activation and lateralization. Similar findings were noted in the contralateral frontoparietal network with decrease in activation relative to peripheral treatment. Other studies have shown normalization of intracortical inhibition after therapeutic BoNT injection (49, 50); however, how these broad changes can generate specific hand muscle movement recovery, as opposed to a more generalized activation, is not readily apparent.
These findings fail to substantiate the proposed mechanism of “activation of dormant central circuitry” controlling the spastic-affected muscles. However, they do support an “unmasking” effect caused by BoNT in the central nervous system. BoNT intervention in dysfunctional musculature generates cortical “normalization” by deactivation of aberrantly stimulated areas (12, 47, 55). A commonly proposed hypothesis for these changes is stabilization of overactive Ia afferent inputs projecting from the spastic muscle by BoNT, in turn, restricting the extent of cortical activation and normalizing motor efferent pathways back into targeted or adjacent muscles (50). In fact, there is experimental evidence that shows how BoNT mediates both extrafusal and intrafusal effects in post-stroke spasticity (56, 57). If BoNT can modulate the intrafusal fibres in the neuromuscular junction, it can decrease overstimulation carried by afferent circuitry that inputs to the cortex. BoNT becomes an agent of partial deafferentation, causing cortical changes, albeit distinct, as the ones observed in multiple studies with distinct pathology. This same mechanism has also been established in post-stroke patients using other means. Sens et al. were able to prove that temporary functional deafferentation by way of an topical anaesthetic cream applied to the forearm resulted in motor and somatosensory improvements in stroke patients, a finding not observed in healthy subjects (58). This is a plausible explanation for muscle recruitment in previously “inactive” hand muscles. That is, by consecutive peripheral BoNT/A injections to the affected hand muscles, the toxin balances the interplay between Ia afferent input and the motor pathway described previously, thus unmasking latent muscle activity.
Most of these studies conclude that BoNT-mediated supraspinal effects are a result of afferent pathway modulation. That is, BoNT injected in the periphery and acting in the neuromuscular junction can reduce input from group Ia afferent fibres. This permits alteration of excitability of spinal pathways as well as motor maps at the cortical level (54). Cortical reorganization after peripheral deafferentation has been well established in other diagnosis, including spinal cord injury (59–62) and amputation, with and without the presence of phantom limb pain (63, 64). Peripheral nerve injury, such as that occurring in brachial plexus injury, again was related to cortical reorganization (65, 66). This has also been demonstrated in healthy individuals after reversible peripheral limb deafferentation experiments (67, 68). As stated, there is sufficient evidence establishing how peripheral deafferentation effects by multiple mechanisms can produce cortical changes, be it in activity or structure. This allows the possibility that a peripheral agent, such as BoNT, can produce cortical changes by means of deafferentation effects, modulating gamma motoneuron endings and reducing group Ia afferent inputs.
Retrograde transport of BoNT through axonal carriers to the spinal cord and subsequent modulation of reciprocal inhibition is another potential explanation of findings in our clinical cases. It has been proposed as a potential mechanism of BoNT-mediated central action by several studies cited previously. Animal models have been utilized to demonstrate this theory. For example, radiolabelled BoNT/A was found in intraspinal motor axons after injection to animal gastrocnemius muscles, indicating retrograde transport of the neurotoxin (69–71). Similar findings were observed in a rat model, where there was a decrease in compound motor action potential amplitudes of the contralateral gastrocnemius after ipsilateral BoNT injection. Of note, inmunohistochemical results revealed BoNT at the bilateral ventral and dorsal horns signalling at a retrograde axonal transport by way of a transcytosis mechanism (72). This was also noted recently using the visual system of a rat model (73). This allows BoNT to act on different structures in the spinal cord, potentially altering the activity of muscles not treated initially. The occurrence of changes in muscle activity and/or parameters distant to BoNT-injected muscle groups has been discussed previously, including modulation of spinal reciprocal inhibition of distant muscles not innervated by the originally injected muscle (9) as well as of antagonist muscles (38, 39). If this is correct, it can provide another potential explanation as to how the patients described in the clinical cases demonstrated motor activity in their spastic-plegic hand muscles after BoNT/A intervention. BoNT could have potentially “disinhibited”, by way of retrograde transport, the antagonist extensor muscles, permitting the patients to actively flex and extend their digits. Yet, the validity of the retrograde transport theory of BoNT in humans has been questioned, which makes this a less likely scenario to explain the unexpected late motor recovery in the clinical cases.
Spastic extremities have been postulated to possess disordered motor control. For instance, the inability to properly extend a spastic joint can be the result of weak extensors due to diminished corticospinal tract activity or tract damage, as well as the co-activation of spastic flexors (74). Impaired motor control can also be explained by dystonia, another manifestation of upper motor neurone syndrome and a variant of muscle overactivity, which can co-exist along with spasticity. Dystonia leads to sustained agonist-antagonist muscle co-contractions and an excess of unwanted movements. It can also lead to abnormal posturing and, eventually, contractures (75). Clinically, dystonic movement may improve with “sensory tricks” (geste antagonistique) (75). For instance, wearing a glove improved motor control in a musician’s dystonia (76). An alteration of the somatosensory input can thus abolish dystonia, albeit, temporarily. BoNT, by way of afferent sensory fibre modulation, could also act as a “sensory trick” acting in the periphery to decrease muscle overactivity. Dystonia is a phenomenon that also has central aberrations. Abnormalities in sensorimotor integration have been reported using somatosensory-evoked potentials and TMS (75). These central alterations observed in dystonia can potentially be modified by the peripheral presence of BoNT acting on the afferent input. Thus, BoNT can act as both a peripheral and, possibly, central modulating agent to improve motor control by acting on multiple permutations of muscle overactivity.
Although the potential of BoNT-mediated central effects after peripheral intervention is exciting, these should not be viewed exclusively as positive outcomes. Weakness of distant muscles after local BoNT intervention can disrupt the kinetic chain of previously adequate muscle group and lead to functional deterioration. Alterations in distant nerve parameters can lead to aberrant or decreased sensation, muscle hyperactivity and others. Not all neural plasticity is beneficial. In fact, maladaptive neural plasticity is prevalent after upper motor neurone injuries and is the target of rehabilitation. Much is unknown about the potential central effects of BoNT. Clinicians should be vigilant of changes in areas distant to the local injection site and evaluate how these could affect the patient’s function.
In the clinical cases reported here, patients could actively flex and extend their previously immobile fingers after BoNT/A injection. A similar case has been reported previously, where BoNT/A injection of spastic finger flexors produced improved voluntary grip control and the ability to open the hand faster (77). Better control of antagonist muscles was also noted in patients with spastic hemiparesis after BoNT injection (78). These findings of late motor recovery cannot be explained solely by spasticity reduction. However, centrally mediated unmasking of antagonist muscles, be it by cortical reorganization after peripheral BoNT-induced deafferentation of gamma motor neurones and/or spinal disinhibition due to retrograde transport of peripherally injected BoNT, are potential mechanisms to explain these favourable outcomes in these clinical cases.
The management of patients with post-stroke spasticity with BoNT has led to observations that cannot be explained exclusively by the peripheral action of the neurotoxin at the neuromuscular junction. The clinical cases briefly described in this review article exemplify late motor recovery noted in patients after peripheral chemodenervation with BoNT. Currently, the most likely mechanism for these observations is the unmasking of latent motor activity in the antagonist muscles by the action of BoNT in the central nervous system accompanied by the reduction in the hyperactivity of the injected agonist muscles. There is a plethora of research demonstrating that this neurotoxin does, in fact, produce actions distant to its peripheral injection site. More so, there is substantial work suggesting that BoNT can generate central modulation by acting on spinal cord neurones and/or alteration of cortical activity, among other potential mechanisms.
Both reported cases arrived at our clinic long after their initial injury (4 years and 38 years later). Although late motor recovery following peripheral BoNT injection is a rare and astounding phenomenon, the case for earlier intervention should be explored. BoNT intervention earlier than 3 months following a stroke has been advocated by experts in the field (79). By injecting earlier rather than later, biomechanical changes that could lead to soft-tissue disorders (i.e. contractures) may be prevented. Early BoNT injection before the actual development of spasticity prevented abnormal posture and contractures in a hereditary spastic mouse model (80). In a recent meta-analysis, Rosales and others (81) reported that early BoNT injection (3 months or earlier) to treat post-stroke spasticity was safe and had a significant treatment effect in most joints studied. Neural plasticity could be fostered by combining early rehabilitation and the centrally-mediated effects of BoNT injection as an adjunct to treat muscle overactivity. By doing so, the occurrence of late maladaptive neuroplasticity could be prevented, such as that seen in these cases.
Numerous questions arise when evaluating the potential of BoNT as a modulating agent of central neural plasticity. If BoNT is to be utilized for a separate therapeutic effect additional to reduction in spasticity, then dosing guidelines must be scrutinized. How often should we inject? How early should we intervene? What amount of BoNT is necessary to produce this beneficial effect? Should this intervention be combined with other treatments, such as physical or occupational therapy? Who will respond favourably with motor recovery after BoNT intervention? Answers to these crucial questions are not readily available. In one of the cases presented, the patient experienced late motor recovery in her hand, without any form of occupational therapy, well within 3 months after intervention. In general, the time between BoNT injections for spasticity is 3 months. These guidelines have been delineated from earlier studies published more than 20 years ago, demonstrating positive effect from injection lasting an average of 10–12 weeks in diagnoses including cervical dystonia (82), spasmodic torticollis (83), hemifacial spasm (84) and other focal dystonias (85). Another reason behind this waiting period between injections is the possibility of producing antibodies to the toxin, thus limiting its effect. However, the BoNT commercial formulations have improved in purity, decreasing the risk of antibody production with repeated injections. For instance, newer formulations have reduced immunogenicity and a lower likelihood for neutralizing antibody production because of decreased protein load, although this fact has yet to be proven in clinical trials (86). Even more so, the risk of producing antibodies to the toxin earlier than expected can be offset by an even earlier resolution of symptoms and restoration of motor activity if more frequent injections are performed. This raises the question: should we inject earlier than 3 months to take advantage of possible central effects of peripheral BoNT injection? Future studies are required to answer these and many other questions regarding the dosing of BoNT intervention for potential central modulation.
In our experience, late motor recovery is not the norm after using BoNT to treat spasticity. Although we hypothesize that a functioning corticospinal tract and other relatively intact central circuitry is necessary for these additional therapeutic effects, more research is required to evaluate the patient demographics where these findings occur, as well as to validate these postulated mechanisms of action. Even though motor recovery occurred in one of our patients without participation in occupational therapy, we must not forget that chemodenervation is but one tool in the physician’s set to treat spasticity. Physical and occupational therapy modalities have been proven to be effective in both the management of spasticity and restoration of motor activity. For instance, the combination of modified constraint-induced therapy and BoNT/A injection to an affected spastic limb has been shown to be efficacious (87). A combination of treatment modalities, including BoNT chemodenervation and therapy, may prove even more beneficial in late motor recovery; however, research is again necessary to corroborate this proposal. It is through this continuous study that BoNT can potentially be extended beyond its status as an effective therapeutic tool against spasticity and into the realm of central-modulating agent, harnessing neuroplasticity in ways that can benefit patients in their daily activities.
There is ample evidence documenting effects caused by peripheral BoNT intervention distant from the primary injection site. These may play a part in clinical results not readily explained by the partial paralysis used to treat spasticity, including late motor recovery. Some of the potential mechanisms behind these beneficial outcomes include: direct action of BoNT at sites in the central nervous system, such as retrograde transport of the neurotoxin into the spinal cord; and cortical reorganization due to peripheral deafferentation at the local injection site. More research is needed to further evaluate these hypotheses as well as the different dosing specifications, patient selection criteria and the interplay with other therapeutic modalities necessary to promote late motor recovery.
The authors declare no conflicts of interest.


 Click to show fullsize
Click to show fullsize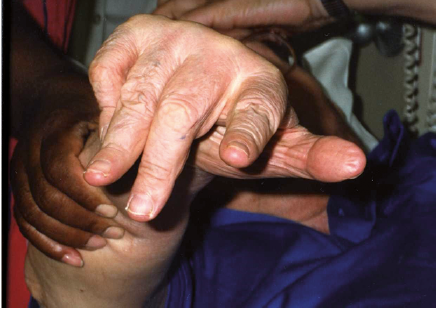 Click to show fullsize
Click to show fullsize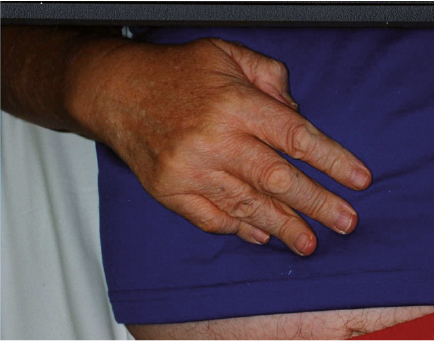 Click to show fullsize
Click to show fullsize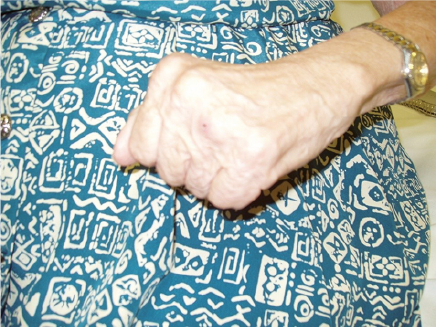 Click to show fullsize
Click to show fullsize